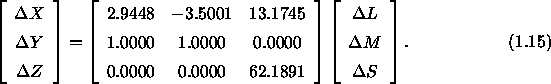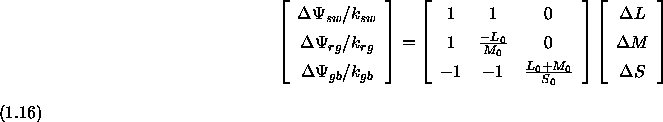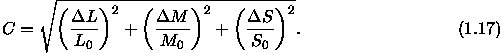Nach der Beschreibung wichtiger Phðnomene der Farbwahrnehmung im vorherigen Abschnitt sollen nun verschiedene Modelle zu deren Erklðrung genannt werden. Wyszecki und Stiles (1982) unterteilen die Theorien zur Farbwahrnehmung in drei Klassen: In diesem Abschnitt sollen die auf den Absorptionsspektren der Rezeptortypen aufbauende Theorien, die neuronalen Theorien, die ■ber die Rezeptorebene hinausgehen, und die Linienelement-Theorien jeweils kurz dargestellt werden.
Die sogenannten Rezeptorkurven-Theorien gehen von den verschiedenen Rezeptortypen auf der Retina aus, die sich in ihrem zu Beginn dieser Arbeit beschriebenen Absorptionsspektrum unterscheiden. Der Absorption liegt das Prinzip der Univarianz zugrunde (Rushton, 1972; Kaiser & Boynton, 1996), das besagt, daÔ zwei beliebige Lichtreize, die vom Sehfarbstoff Rhodopsin gleich absorbiert werden, auch zu einer gleichen Reaktion der Zapfen f■hren. Die f■r jeden Zapfentyp charakteristische spektrale Empfindlichkeit beschreibt die Wahrscheinlichkeit, mit der ein Photon absorbiert wird, als Funktion der Wellenlðnge (siehe Abbildung 1.1). Die aus der Absorption resultierende Zapfenerregung hðngt nur von der Anzahl absorbierter Photonen ab, nicht von deren Wellenlðnge, da die Wellenlðngeninformation verloren geht, sobald ein Photon erst einmal absorbiert worden ist. Aus dem Prinzip der Univarianz folgt, daÔ die spektralen Hellempfindlichkeitskurven der Zapfentypen linear mit den relativen Absorptionsspektren der drei Farbrezeptor-Arten zusammenhðngen m■ssen (siehe Wyszecki & Stiles, 1982, S. 587). Diese spektralen Empfindlichkeiten nennt man auch K—nig-Fundamentals nach A. K—nig, von dem die ersten empirisch ermittelten Kurven dieser Art stammen.
Diese K—nig-Fundamentals stimmen gut mit den spektralen Empfindlichkeiten der ![]() -Mechanismen
-Mechanismen![]() von Stiles (1959) ■berein und auch die von Baylor et al. (1987) und Schnapf et al. (1987) berichteten Absorptionsspektren der verschiedenen Pigmente der Zapfentypen sind ebenfalls sehr ðhnlich. Verschiedenste Arten, diese fundamentalen Empfindlichkeitsfunktionen der Zapfen zu bestimmen, f■hren also zu sehr ðhnlichen Ergebnissen, deren geringf■gige Unterschiede von Wyszecki und Stiles (1982, S. 633) diskutiert werden. Das Besondere an dieser Ébereinstimmung ist, daÔ die Daten auf v—llig verschiedene Weise zustande kommen: Die K—nig-Fundamentals basieren auf Farbwertkurven und Verwechslungslinien, also letztlich auf Farbabgleichen, die Empfindlichkeiten der
von Stiles (1959) ■berein und auch die von Baylor et al. (1987) und Schnapf et al. (1987) berichteten Absorptionsspektren der verschiedenen Pigmente der Zapfentypen sind ebenfalls sehr ðhnlich. Verschiedenste Arten, diese fundamentalen Empfindlichkeitsfunktionen der Zapfen zu bestimmen, f■hren also zu sehr ðhnlichen Ergebnissen, deren geringf■gige Unterschiede von Wyszecki und Stiles (1982, S. 633) diskutiert werden. Das Besondere an dieser Ébereinstimmung ist, daÔ die Daten auf v—llig verschiedene Weise zustande kommen: Die K—nig-Fundamentals basieren auf Farbwertkurven und Verwechslungslinien, also letztlich auf Farbabgleichen, die Empfindlichkeiten der ![]() -Mechanismen werden aus Schwellen f■r wahrnehmbare Inkremente abgeleitet und die Absorptionskurven der Sehpigmente werden durch physiologische Messungen bestimmt. Diese drei unterschiedlichen Herangehensweisen belegen alle, daÔ drei Mechanismen mit einer bestimmten Charakteristik die Grundlage des Farbensehens bilden.
-Mechanismen werden aus Schwellen f■r wahrnehmbare Inkremente abgeleitet und die Absorptionskurven der Sehpigmente werden durch physiologische Messungen bestimmt. Diese drei unterschiedlichen Herangehensweisen belegen alle, daÔ drei Mechanismen mit einer bestimmten Charakteristik die Grundlage des Farbensehens bilden.
Rushton (1972) schlðgt ein Modell der Farbwahrnehmung vor, das direkt auf den Reaktionen der Rezeptortypen basiert und das Farbadaptation dadurch erklðrt, daÔ die Pigmente der Rezeptoren bei Absorption von Licht bleichen, d.h. in einen anderen chemischen Zustand zerfallen, wodurch sich die Empfindlichkeit der Zapfensysteme verðndert (siehe auch Kaiser & Boynton, 1996). Dabei herrscht ein chemisches Gleichgewicht zwischen der gebleichten und der ungebleichten Form des Sehpigments, das mit zunehmender Absorption von Photonen in Richtung gebleichter Pigmente verschoben wird. Dadurch wird eine verstðrkte Neigung zum Wiederherstellen der gebleichten Molek■le ausgel—st.
MacLeod und Boynton (1979) stellen einen dreidimensionalen Farbraum vor, der direkt auf physiologischen Messungen aufbaut und dessen Achsen durch die Erregung der drei Rezeptortypen L, M und S gegeben sind. Sie gehen von der Annahme aus, daÔ die Rezeptoren des S-Systems nicht zur Gesamt-Leuchtdichte beitragen und daÔ die Summe der Erregungen von L und M konstant sei und damit gleich Eins gesetzt werden kann. Dann lassen sich die Koordinaten einer Farbe in diesem Raum folgendermaÔen reprðsentieren:
![]()
MacLeod und Boynton (1979) nennen als Vorteile dieser Darstellungsweise, daÔ die Erregung der einzelnen Rezeptorsysteme direkt reprðsentiert ist und daÔ gleichen Entfernungen auf einer Linie in dieser Farbtafel auch gleiche Verðnderungen der Rezeptorerregungen f■r die drei Zapfenarten entsprechen. Dieser Farbraum dient als Grundlage f■r den in Abschnitt 1.2.3 beschriebenen DKL-Farbraum.
Das farbliche Aussehen eines Reizes wird nicht nur von der Erregung der drei Rezeptorarten bestimmt, sondern auch durch eine Weiterverarbeitung der Rezeptorsignale auf h—herer neuronaler Ebene, wie unter anderem von Jameson und Hurvich (1964) oder Shevell (1978, 1982) belegt wird. Alle derartigen Modelle, die sogenannten Gegenfarben-Theorien, gehen dabei von antagonistischen Farbmechanismen auf dieser h—heren Ebene aus. Hering (1920) postuliert beispielsweise drei antagonistische Prozesse als Grundlage des Farbensehens, nðmlich einen Rot-Gr■n-ProzeÔ, einen Gelb-Blau-ProzeÔ und einen WeiÔ-Schwarz-ProzeÔ, wobei keine Farbe gleichzeitig beide Farbt—ne des jeweiligen Antagonisten-Paares annehmen kann.
Jameson und Hurvich (1955) quantifizieren dieses Modell durch Anwendung der Farbaufhebungs-Methode (color cancellation method), bei der das Mischungsverhðltnis von zwei Farbreizen so eingestellt werden soll, daÔ deren Mischung weder r—tlich noch gr■nlich (bzw. weder blðulich noch gelblich) erscheint. Der Rotanteil eines Reizes lðÔt sich dann dadurch bestimmen, wieviel Gr■n hinzugemischt werden muÔ, um eine weder r—tliche noch gr■nliche Mischung zu erhalten. Die Erregung des Rot-Gr■n-Mechanismus durch einen Spektralreiz der Wellenlðnge ![]() lðÔt sich durch die Differenz zwischen der Intensitðt des roten Anteils und des gr■nen Anteils
lðÔt sich durch die Differenz zwischen der Intensitðt des roten Anteils und des gr■nen Anteils ![]() beschreiben; ðhnlich verhðlt es sich mit der Intensitðtsdifferenz
beschreiben; ðhnlich verhðlt es sich mit der Intensitðtsdifferenz ![]() des Gelb-Blau-Systems. Durch Variieren der Wellenlðnge
des Gelb-Blau-Systems. Durch Variieren der Wellenlðnge ![]() lassen sich so die spektralen Reaktionskurven der Gegenfarben-Systeme empirisch bestimmen. Hurvich und Jameson (1955, S.602) geben aber auch an, wie diese Reaktionskurven als Linearkombination der CIE-1931-Spektralwertkurven berechnet werden k—nnen (empirische Befunde, die gegen diese Linearitðtsannahme sprechen, werden im nðchsten Abschnitt besprochen). Jameson & Hurvich (1956) beschreiben, wie auch Kontexte und neutrale Reize im Rahmen dieser Theorie ber■cksichtigt werden k—nnen.
lassen sich so die spektralen Reaktionskurven der Gegenfarben-Systeme empirisch bestimmen. Hurvich und Jameson (1955, S.602) geben aber auch an, wie diese Reaktionskurven als Linearkombination der CIE-1931-Spektralwertkurven berechnet werden k—nnen (empirische Befunde, die gegen diese Linearitðtsannahme sprechen, werden im nðchsten Abschnitt besprochen). Jameson & Hurvich (1956) beschreiben, wie auch Kontexte und neutrale Reize im Rahmen dieser Theorie ber■cksichtigt werden k—nnen.
Die von Jameson und Hurvich (1955, 1956) und Hurvich und Jameson (1955) dargestellte Gegenfarben-Theorie dient zur Beschreibung der Ergebnisse von Farbaufhebungs-Experimenten, die formalen Voraussetzungen f■r eine Gegenfarben-Theorie werden aber von diesen Autoren nicht betrachtet. Von Krantz (1975b) wird dazu die Gegenfarben-Theorie axiomatisiert, indem die aus Abschnitt 1.1.2 bekannte GraÔmann-Struktur ![]() um die Teilmengen
um die Teilmengen ![]() (Rot-Gr■n-áquilibria) und
(Rot-Gr■n-áquilibria) und ![]() (Gelb-Blau-áquilibria), zu einer Gegenfarben-Struktur
(Gelb-Blau-áquilibria), zu einer Gegenfarben-Struktur ![]() erweitert wird. Sind neben den f■r eine trichromatische GraÔmann-Struktur notwendigen Axiomen noch drei Axiome empirisch erf■llt, die die Linearitðt der Gegenfarben-Attribute sicherstellen, dann gilt der folgende Reprðsentationssatz (Theorem 6 bei Krantz, 1975b):
erweitert wird. Sind neben den f■r eine trichromatische GraÔmann-Struktur notwendigen Axiomen noch drei Axiome empirisch erf■llt, die die Linearitðt der Gegenfarben-Attribute sicherstellen, dann gilt der folgende Reprðsentationssatz (Theorem 6 bei Krantz, 1975b):
Dies bedeutet, daÔ die Funktionen ![]() und
und ![]() eindeutig sind bis auf MaÔstabsðnderungen und Vorzeichenwechsel.
eindeutig sind bis auf MaÔstabsðnderungen und Vorzeichenwechsel.
Krantz (1975b) zeigt auÔerdem, daÔ auch dann ein linearer Farbcode auf der GraÔmann-Struktur konstruiert werden kann, wenn das Axiom der Linearitðt nur von einer einzigen áquilibriums-Menge erf■llt wird. Dies ist aus empirischen Gr■nden von groÔer Bedeutung, denn Larimer, Krantz und Cicerone (1974) k—nnen zwar f■r die Rot-Gr■n-Koeffizienten keine Verletzung der Linearitðt nachweisen, aber f■r die Gelb-Blau-Koeffizienten weisen Larimer, Krantz und Cicerone (1975) deutliche Abweichungen von der Linearitðt nach. Ikeda und Ayama (1980) k—nnen unter Anwendung der oben beschriebenen Farbaufhebungs-Methode Hinweise darauf finden, daÔ die Gelbempfindung aus einer nichtlinearen Kombination zweier Zapfenreaktionen resultiert. Mausfeld (1998) greift die Axiomatisierung der Gegenfarben-Theorie von Krantz (1975b) auf und nennt weitere empirische Belege f■r Verletzungen der geforderten Axiome.
Trotz dieser Verletzungen der Linearitðt soll nun ein auf Gegenfarben aufbauender Farbraum dargestellt werden, der eine anschauliche Reprðsentation von Innenfeld-Umfeld-Konfigurationen erlaubt, nðmlich der von Derrington, Krauskopf und Lennie (1984) entwickelte und nach ihnen benannten DKL-Farbraum, dem die auf Seite ![]() beschriebene auf den Rezeptorerregungen der L-, M- und S-Zapfen aufbauende Farbtafel von MacLeod und Boynton (1979) zugrunde liegt.
beschriebene auf den Rezeptorerregungen der L-, M- und S-Zapfen aufbauende Farbtafel von MacLeod und Boynton (1979) zugrunde liegt.
Derrington et al. (1984) postulieren aufgrund von Versuchen an der Retina von Makaken die Existenz von drei antagonistischen neuronalen Mechanismen, die die Achsen ihres Farbraums bilden: Eine chromatische Dimension, deren Ausprðgung sich aus den Zapfenerregungen als S-(L+M) berechnet, wird als konstante S-Erregung ![]() bezeichnet und entspricht dem Gelb-Blau-Mechanismus. Der zweite chromatische Gegenfarben-Mechanismus, dessen Aktivierung als Differenz der Zapfenerregungen L-M bestimmt wird, nennt sich konstante L-M-Erregung
bezeichnet und entspricht dem Gelb-Blau-Mechanismus. Der zweite chromatische Gegenfarben-Mechanismus, dessen Aktivierung als Differenz der Zapfenerregungen L-M bestimmt wird, nennt sich konstante L-M-Erregung ![]() und entspricht dem Rot-Gr■n-Mechanismus. Hinzu kommt noch eine dritte Achse
und entspricht dem Rot-Gr■n-Mechanismus. Hinzu kommt noch eine dritte Achse ![]() , die die Leuchtdichte-Information enthðlt. Dieser achromatische Mechanismus hðngt nicht von der Erregung des S-Systems ab und errechnet sich als Summe der Zapfenerregungen L+M. Diese drei antagonistischen Mechanismen bilden die Grundlage von rezeptiven Feldern. Das Modell von Derrington et al. (1984) postuliert auÔerdem eine Normierung zum WeiÔpunkt
, die die Leuchtdichte-Information enthðlt. Dieser achromatische Mechanismus hðngt nicht von der Erregung des S-Systems ab und errechnet sich als Summe der Zapfenerregungen L+M. Diese drei antagonistischen Mechanismen bilden die Grundlage von rezeptiven Feldern. Das Modell von Derrington et al. (1984) postuliert auÔerdem eine Normierung zum WeiÔpunkt ![]() , der entweder dem energiegleichen Spektrum oder dem Farbort des Kontextes entspricht und als Ursprung des Gegenfarben-Koordinatensystems gewðhlt wird. Als Farbkoordinaten eines Reizes, die nun als DKL-Koordinaten bezeichnet werden, werden die Differenzen zu diesem Punkt auf den drei gerade beschriebenen Achsen eingesetzt. Die genaue Berechnung dieser DKL-Koordinaten wird im nðchsten Abschnitt dargestellt.
, der entweder dem energiegleichen Spektrum oder dem Farbort des Kontextes entspricht und als Ursprung des Gegenfarben-Koordinatensystems gewðhlt wird. Als Farbkoordinaten eines Reizes, die nun als DKL-Koordinaten bezeichnet werden, werden die Differenzen zu diesem Punkt auf den drei gerade beschriebenen Achsen eingesetzt. Die genaue Berechnung dieser DKL-Koordinaten wird im nðchsten Abschnitt dargestellt.
Die Bestimmung der DKL-Farbkoordinaten aus den Normfarbwerten eines Reizes und seines Umfelds erfordert mehrere Rechenschritte, die nun entsprechend Brainard (1996) dargestellt werden. Den Ausgangspunkt bilden die Differenzen ![]() ,
, ![]() und
und ![]() zwischen den von Reiz und Kontext hervorgerufenen Rezeptorerregungen, die nach Brainard (1996) mit den Differenzen
zwischen den von Reiz und Kontext hervorgerufenen Rezeptorerregungen, die nach Brainard (1996) mit den Differenzen ![]() ,
, ![]() und
und ![]() der Normfarbwerte von Reiz und Kontext folgendermaÔen zusammenhðngen:
der Normfarbwerte von Reiz und Kontext folgendermaÔen zusammenhðngen:
Aus der Gleichung 1.15 lassen sich Vorhersagen ■ber die Aktivierung der hier als ![]() bezeichneten drei Mechanismen berechnen, wobei die Konstanten
bezeichneten drei Mechanismen berechnen, wobei die Konstanten ![]() ,
, ![]() und
und ![]() die Einheit des jeweiligen Reaktionssystems festlegen: Die Erregung des achromatischen Mechanismus
die Einheit des jeweiligen Reaktionssystems festlegen: Die Erregung des achromatischen Mechanismus ![]() berechnet sich nach der zweiten Zeile von Gleichung 1.15, die die mit der Leuchtdichtedifferenz gleichgesetzte Erregungsdifferenz
berechnet sich nach der zweiten Zeile von Gleichung 1.15, die die mit der Leuchtdichtedifferenz gleichgesetzte Erregungsdifferenz ![]() angibt:
angibt:
![]()
F■r den L-M-Mechanismus ![]() wird nach Brainard (1996) hier eine gewichtete Differenz
wird nach Brainard (1996) hier eine gewichtete Differenz ![]() bestimmt, die auch vom Hintergrund abhðngt, wobei
bestimmt, die auch vom Hintergrund abhðngt, wobei ![]() eine gewichtete Summe aus der Erregungsdifferenz zwischen L- und M-System und der Erregung des L-Systems bezeichnet:
eine gewichtete Summe aus der Erregungsdifferenz zwischen L- und M-System und der Erregung des L-Systems bezeichnet:
![]()
Die Aktivierung des S-Mechanismus ![]() hðngt nach Brainard (1996) ebenfalls vom Kontext ab.
hðngt nach Brainard (1996) ebenfalls vom Kontext ab. ![]() ist eine gewichtete Summe aus der Erregungsdifferenz zwischen S- und achromatischem System und der Erregung des S-Systems:
ist eine gewichtete Summe aus der Erregungsdifferenz zwischen S- und achromatischem System und der Erregung des S-Systems:
![]()
Aufgrund dieser Vor■berlegungen k—nnen nun aus den Zapfenerregungs-Differenzen ![]() und
und ![]() die Reaktionen der drei Gegenfarben-Mechanismen berechnet werden. Brainard (1996) setzt dazu
die Reaktionen der drei Gegenfarben-Mechanismen berechnet werden. Brainard (1996) setzt dazu ![]() und
und ![]() und gelangt so zu folgender Transformation vom Raum der Rezeptorerregungen in den DKL-Raum:
und gelangt so zu folgender Transformation vom Raum der Rezeptorerregungen in den DKL-Raum:
Er empfiehlt, ![]() so zu wðhlen, daÔ
so zu wðhlen, daÔ ![]() dem Helligkeitskontrast C entspricht, der beispielsweise aus den Kontrasten der Zapfenerregungen
dem Helligkeitskontrast C entspricht, der beispielsweise aus den Kontrasten der Zapfenerregungen ![]() ,
, ![]() und
und ![]() berechnet werden kann (diese und andere Varianten werden von Brainard (1996, S. 568) diskutiert):
berechnet werden kann (diese und andere Varianten werden von Brainard (1996, S. 568) diskutiert):
Derrington et al. (1984) wðhlen die Werte von ![]() und
und ![]() so, daÔ die maximal in ihrem Versuchsaufbau realisierbaren Erregungen
so, daÔ die maximal in ihrem Versuchsaufbau realisierbaren Erregungen ![]() und
und ![]() den Wert 1.0 erhalten. Brainard (1996) schlðgt eine andere M—glichkeit zur Bestimmung der Skalierungsfaktoren f■r die drei Achsen vor: Er normiert die drei Mechanismen durch geeignete Wahl der Konstanten
den Wert 1.0 erhalten. Brainard (1996) schlðgt eine andere M—glichkeit zur Bestimmung der Skalierungsfaktoren f■r die drei Achsen vor: Er normiert die drei Mechanismen durch geeignete Wahl der Konstanten ![]() und
und ![]() so, daÔ sich bei isolierter Erregung eines der drei Systeme durch einen Reiz mit einem Helligkeitskontrast von einer Einheit jeweils der Wert Eins f■r die Reaktion des Gegenfarben-Systems ergibt. Nach Gleichung 1.16 gelangt man dabei zu folgenden Formeln zur Bestimmung dieser Normierungsfaktoren:
so, daÔ sich bei isolierter Erregung eines der drei Systeme durch einen Reiz mit einem Helligkeitskontrast von einer Einheit jeweils der Wert Eins f■r die Reaktion des Gegenfarben-Systems ergibt. Nach Gleichung 1.16 gelangt man dabei zu folgenden Formeln zur Bestimmung dieser Normierungsfaktoren:
![]()
Die Differenzen der Erregung der antagonistischen Systeme lassen sich damit aus den Differenzen der Rezeptorerregung nach der folgenden Gleichung berechnen:
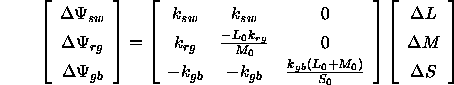
Die Erregungsdifferenzen der Gegenfarben-Kanðle lassen sich auch direkt durch Anwenden von Gleichung 1.15 aus den Normfarbwerten von Reiz und Hintergrund bestimmen. An diesem Farbmodell kann kritisiert werden, daÔ die Berechnung der Skalierungsfaktoren ![]() ,
, ![]() und
und ![]() nicht aus theoretischen Gr■nden erfolgt, sondern nur aus Plausibilitðtsgr■nden auf die hier beschriebene Weise stattfindet. Bei der Interpretation der Einheiten der Achsen in diesem Farbraum sollte dies immer ber■cksichtigt werden. áhnliches gilt f■r die Berechnung des Helligkeitskontrastes, der ja ebenfalls an der Normierung der Achsen beteiligt ist.
nicht aus theoretischen Gr■nden erfolgt, sondern nur aus Plausibilitðtsgr■nden auf die hier beschriebene Weise stattfindet. Bei der Interpretation der Einheiten der Achsen in diesem Farbraum sollte dies immer ber■cksichtigt werden. áhnliches gilt f■r die Berechnung des Helligkeitskontrastes, der ja ebenfalls an der Normierung der Achsen beteiligt ist.
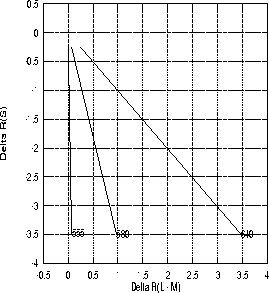
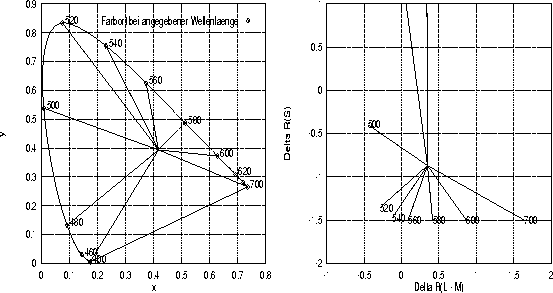
Gleiche Farbart bei unterschiedlicher Leuchtdichte im DKL-Raum: Hier ist die Erregungsdifferenz der beiden chromatischen Gegenfarben-Systeme f■r die drei im Experiment verwendeten monochromatischen Reize (Wellenlðnge 555, 580 und 640 nm) im DKL-Raum eingetragen. Jede der drei Linien beginnt bei einer Leuchtdichte des monochromatischen Reizes von 0.5 ![]() und endet bei 7.0
und endet bei 7.0 ![]() . Den Berechnungen wird als Umfeld der auch im Experiment verwendete Ausgangskontext mit den xyL-Koordinaten (0.25, 0.35, 2.0) zugrundegelegt.
. Den Berechnungen wird als Umfeld der auch im Experiment verwendete Ausgangskontext mit den xyL-Koordinaten (0.25, 0.35, 2.0) zugrundegelegt.
Betrachtet man im DKL-Raum einen Reiz vor einem festen Kontext und variiert nur dessen Leuchtdichte, dann liegen alle diese Variationen auf einer Gerade, die auÔerdem durch den Ursprung geht, wie an dem Beispiel in Abbildung 1.5 zu sehen ist. Ein derartiges Muster in den bei Farbabgleichen erhobenen Daten w■rde damit bedeuten, daÔ die Verhðltnisse der Erregungsdifferenzen genau repliziert werden k—nnen, wobei allerdings die Leuchtdichte der Einstellungen stðrkeren Schwankungen unterliegt. Wie man in Abbildung 1.6 sehen kann, liegen bunttongleiche Farben, die sich nur in ihrer Sðttigung unterscheiden, sowohl in der Normfarbtafel als auch im DKL-Raum alle auf einer Strecke, deren Endpunkte die Farbvalenzen des monochromatischen Reizes und das hinzugemischten ``WeiÔ'' bilden. Auch Sðttigungsunterschiede f■hren also im DKL-Raum zu Koordinaten, die auf einer Geraden liegen; diese Geraden treffen sich f■r verschiedene Reize jeweils im WeiÔpunkt.
Bunttongleiche Farben in der Normfarbtafel und im DKL-Raum: Diese Abbildung zeigt die Farborte in der Normfarbtafel (links) und im DKL-Raum (rechts) von verschiedenen monochromatischen Reizen sowie die Strecken, auf denen die Mischungen dieser monochromatischen Reize mit einem ``weiÔen'' Reiz liegen, der dem im Experiment verwendeten Entsðttigungsreiz mit den Koordinaten x= 0.4170 und y = 0.3935 entspricht. Sowohl f■r den Kontext als auch f■r den Zielreiz wird dabei von einer konstanten Leuchtdichte von 3 ![]() ausgegangen.
ausgegangen.
Der DKL-Raum der Gegenfarben eignet sich zur Reprðsentation von Farben besonders gut, wenn deren Kontextabhðngigkeit ber■cksichtigt werden soll. Die Darstellung dieses Farbraums durch Brainard (1996) umfaÔt alle f■r die Bestimmung der DKL-Koordinaten erforderlichen Rechenschritte. Diese Theorie ist dadurch so genau spezifiziert, daÔ sie sich auch direkt zur Untersuchung von Kontexteffekten anwenden lðÔt.
Den aus Rezeptorkurven- und Gegenfarben-Theorien entwickelten Farbrðumen, die in den letzten Abschnitten beschrieben sind, liegen letztlich spektrale Empfindlichkeitskurven zugrunde, die alleine aus Urteilen ■ber die Gleichheit von Farben bestimmt werden k—nnen. Deshalb lassen sich aus diesen Theorien keine Aussagen ■ber die Gr—Ôe von Farbunterschieden ableiten. Die Linienelement-Theorien sind der Versuch einer solchen h—heren Farbmetrik, d.h. einer Farbmetrik, die nicht nur Gleichheit bzw. Verschiedenheit von Farben behandelt, sondern Farben so quantifiziert, daÔ sich ein numerisches MaÔ f■r Farbunterschiede bestimmen lðÔt. Empirische Basis sind nicht mehr - wie bei der niederen Farbmetrik - Urteile ■ber die Gleichheit bzw. Verschiedenheit von Farben, sondern ebenmerkliche Farbunterschiede.
Die Grundidee der Linienelement-Theorien im Farbraum geht auf Helmholtz (1891) zur■ck, der davon ausgeht, daÔ die Unterschiedsschwelle der drei Farbsysteme nach dem Weberschen Gesetz von deren Erregungsniveau abhðngt und f■r mittlere Leuchtdichte-Niveaus etwa konstant ist. Dann lðÔt sich ein ebenmerklicher Farb-Unterschied (bzw. das Linienelement d s) wie folgt berechnen:
Die nach diesem Modell errechneten Spektralwertkurven ![]() stimmen jedoch nicht mit den heute bekannten Absorptionsspektren der drei Farbpigmente ■berein. Schr—dinger (1920) versucht die Unzulðnglichkeiten der Helmholtzschen Theorie durch folgende ánderung auszubessern:
stimmen jedoch nicht mit den heute bekannten Absorptionsspektren der drei Farbpigmente ■berein. Schr—dinger (1920) versucht die Unzulðnglichkeiten der Helmholtzschen Theorie durch folgende ánderung auszubessern:
![]()
wobei ![]() ,
, ![]() und
und ![]() nicht nðher spezifizierte Farbkoordinaten sein sollen. Schr—dinger legt seinen Berechnungen die K—nig-Fundamentals als spektrale Empfindlichkeitskurven zugrunde. Bei geeigneten Schðtzwerten f■r die metrischen Koeffizienten
nicht nðher spezifizierte Farbkoordinaten sein sollen. Schr—dinger legt seinen Berechnungen die K—nig-Fundamentals als spektrale Empfindlichkeitskurven zugrunde. Bei geeigneten Schðtzwerten f■r die metrischen Koeffizienten ![]() stimmen die vom daraus bestimmten Linienelement vorhergesagten Hellempfindlichkeiten und die empirisch ermittelte Hellempfindlichkeitsfunktion
stimmen die vom daraus bestimmten Linienelement vorhergesagten Hellempfindlichkeiten und die empirisch ermittelte Hellempfindlichkeitsfunktion ![]() gut ■berein.
gut ■berein.
Stiles (1946) modifiziert die Helmholtzsche Linienelement-Theorie ebenfalls, aber nicht so drastisch wie Schr—dinger: Aufgrund der von ihm empirisch belegten Tatsache, daÔ f■r die drei visuellen Mechanismen unterschiedliche Weber-Br■che vorliegen, postuliert Stiles (1946) aufbauend auf empirisch bestimmten Farbwertkurven ![]() folgendes Linienelement:
folgendes Linienelement:
wobei f■r den von ihm untersuchten Beobachter ![]() ,
, ![]() und
und ![]() gilt. Bei hohen Leuchtdichten reduziert sich das in Gleichung 1.19 definierte Linienelement zu folgender, dem Helmholtzschen Linienelement in Gleichung 1.18 ðhnlichen Form:
gilt. Bei hohen Leuchtdichten reduziert sich das in Gleichung 1.19 definierte Linienelement zu folgender, dem Helmholtzschen Linienelement in Gleichung 1.18 ðhnlichen Form:
![]()
Dieses Linienelement sagt einige empirische Befunde gut vorher, beispielsweise ebenmerkliche Wellenlðngenunterschiede, spektrale Hellempfindlichkeiten und Farbunterschiedsschwellen in Form von MacAdam-Ellipsen. Zwischen den unter anderem von MacAdam (1942b) experimentell bestimmten Diskriminationsellipsen und den von diesem Linienelement vorhergesagten ergeben sich aber prinzipielle Unterschiede: Die GauÔsche Kr■mmung (siehe Gray, 1993) des von Stiles (1946) vorhergesagten Linienelementes ist immer negativ, wðhrend die empirisch bestimmte manchmal negativ und manchmal positiv ist. Es lðÔt sich also prinzipiell keine isometrische Transformation finden, die die empirische und die aus dem Linienelement vorhergesagte Oberflðche gleichen Farbabstandes ineinander ■berf■hrt (siehe hierzu auch Wyszecki & Stiles, 1982).
Trabka (1968a, 1968b) wendet die Konzepte der Signalentdeckungstheorie auf das Linienelement von Stiles (1946) an. Er betrachtet die einzelnen Zapfen als nicht-ideale Empfðnger elektromagnetischer Schwingungen und alle Rezeptoren eines Typs zusammen als einen Detektor-Mechanismus. AuÔerdem nimmt Trabka (1968a) an, daÔ mit dem Signal-Rausch-Abstand auch die Verðnderung des Outputs der Detektor-Mechanismen zunimmt und somit auch der wahrgenommene Abstand zwischen den zwei Farben. Das Linienelement berechnet sich dann als Summe des Signal-Rausch-Abstands am Output von verschiedenen Rezeptoren, die sich nur in ihrer spektralen Absorptionsfunktion und in ihrem Verstðrkungsmechanismus unterscheiden.
Von Vos und Walraven (1972a, 1972b) stammt die komplexeste Linienelement-Theorie, die auf dem Konzept der Gegenfarben aufbaut. Sie nehmen an, daÔ Farbunterscheidbarkeit durch Variabilitðt in der Absorption von Photonen begrenzt wird und daÔ bei h—heren Leuchtdichten Sðttigungs- bzw. Ébersðttigungs-Phðnomene![]() auftreten k—nnen, die dazu f■hren, daÔ das Farbdiskriminationsverm—gen nicht so hoch ist wie alleine aufgrund der Varianz in den Farbkanðlen zu erwarten wðre. Die Signale der Gegenfarben-Kanðle vereinigen sich in einer positiv definiten quadratischen Form zu einem Output-Signal, das direkt proportional zum ebenmerklichen Unterschied zwischen zwei Reizen ist. Mit diesem Modell lassen sich Daten zur Farbdiskrimination recht gut vorhersagen, sowohl was die Wellenlðngen-Diskrimination betrifft als auch bez■glich der Farbton-Unterscheidbarkeit.
auftreten k—nnen, die dazu f■hren, daÔ das Farbdiskriminationsverm—gen nicht so hoch ist wie alleine aufgrund der Varianz in den Farbkanðlen zu erwarten wðre. Die Signale der Gegenfarben-Kanðle vereinigen sich in einer positiv definiten quadratischen Form zu einem Output-Signal, das direkt proportional zum ebenmerklichen Unterschied zwischen zwei Reizen ist. Mit diesem Modell lassen sich Daten zur Farbdiskrimination recht gut vorhersagen, sowohl was die Wellenlðngen-Diskrimination betrifft als auch bez■glich der Farbton-Unterscheidbarkeit.
Einige der hier vorgestellten Linienelement-Theorien k—nnen das Farbdiskriminationsverm—gen relativ gut vorhersagen. Dadurch kann ein Farbraum so transformiert werden, daÔ gleiche Abstðnde in diesem Farbraum auch gleiche Farbunterschiede bedeuten. Wandell (1985) weist allerdings darauf hin, daÔ dies nur f■r konstante Beobachtungsbedingungen gilt, da die metrischen Koeffizienten des Linienelements vom Adaptationszustand des Auges abhðngig sind. Wird dieser aber konstant gehalten, dann lðÔt sich die Unterscheidbarkeit zweier Reize alleine aus der Differenz der die beiden Reize reprðsentierenden Vektoren bestimmen. Wandell (1982, 1985) bezeichnet diese Éberlegung als Vektordifferenz-Hypothese und betrachtet sie als Spezialfall des Linienelements.
Zur Vorhersage der Ergebnisse in dem hier zu beschreibenden Experiment sind die Linienelement-Theorien nur bedingt geeignet, da sie eben keine Kontexteffekte ber■cksichtigen. AuÔerdem lassen sich aus diesen Theorien keine Vorhersagen ■ber m—gliche Auswirkungen der spðter ausf■hrlich diskutierten Gedðchtnismethode ableiten.